

Molecular Evidence Reveals Taxonomic Uncertainties and Cryptic Diversity in the Neotropical Catfish of the Genus Pimelodus (Siluriformes: Pimelodidae)
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Extraction, Amplification, and Sequencing of the DNA
2.3. The DNA Barcoding Analysis and Delimitation of the MOTUs
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Species by Family/Subfamily. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp (accessed on 14 November 2023).
- Teugels, G.G. Taxonomy, phylogeny and biogeography of catfishes (Ostariophysi, Siluroidei): An overview. Aquat. Living Resour. 1996, 9, 9–34. [Google Scholar] [CrossRef]
- Rocha, M.S. Family Pimelodidae—Long-whiskered catfishes. In Field Guide to the Fishes of the Amazon, Orinoco, and Guianas; Van Der Sleen, P., Albert, J.S., Eds.; Princeton University Press: Princeton, NJ, USA, 2018; pp. 299–308. ISBN 978-0-691-17074-9. [Google Scholar]
- Lundberg, J.G.; Littmann, M.W. Family Pimelodidae (Long-whiskered catfishes). In Check List of the Freshwater Fishes of South and Central America; Reis, R.E., Kullander, S.O., Ferraris, C.J., Jr., Eds.; Edipucrs: Porto Alegre, Brazil, 2003; pp. 432–446. [Google Scholar]
- Villa-Navarro, F.A.; Acero, P.A.; Cala, P.C. Taxonomic review of Trans-Andean species of Pimelodus (Siluriformes: Pimelodidae), with the descriptions of two new species. Zootaxa 2017, 4299, 337–360. [Google Scholar] [CrossRef]
- Silva, C.T.; Ribeiro, F.V.R.; Lucena, C.A.S.; Lucinda, P.H.F. Pimelodus speciosus (Teleostei: Pimelodidae), a new catfish species from the rio Tocantins drainage, Brazil. Ichthyol. Explor. Freshw. 2018, 28, 97–106. [Google Scholar]
- Lundberg, J.G.; Sullivan, J.P.; Hardman, M. Phylogenetics of the South American catfish family Pimelodidae (Teleostei: Siluriformes) using nuclear and mitochondrial gene sequences. Proc. Acad. Nat. Sci. Phila. 2011, 161, 153–189. [Google Scholar] [CrossRef]
- Lundberg, J.G.; Covain, R.; Sullivan, J.P.; Fisch-Muller, S. Phylogenetic position and notes on the natural history of Pimelabditus moli Parisi & Lundberg, 2009 (Teleostei: Siluriformes), a recently discovered pimelodid catfish from the Maroni River basin. Cybium 2012, 36, 105–114. [Google Scholar]
- Souza-Filho, H.S.; Shibatta, A.O. Descrição de uma espécie nova de Pimelodus (Siluriformes, Pimelodidae) da bacia do alto rio Paraguai. Iheringia Sér. Zool 2007, 97, 472–480. [Google Scholar] [CrossRef]
- Lundberg, J.G.; Parisi, B.M. Propimelodus, new genus, and redescription of Pimelodus eigenmanni Van der Stigchel 1946, a long-recognized yet poorly-known South American catfish (Pimelodidae: Siluriformes). Proc. Acad. Nat. Sci. Phila. 2002, 152, 75–88. [Google Scholar] [CrossRef]
- Ribeiro, F.R.V.; Lucena, C.A.S. A new species of Pimelodus La Cépède, 1803 (Siluriformes: Pimelodidae) from the rio São Francisco drainage, Brazil. Neotrop. Ichthyol. 2006, 4, 411–418. [Google Scholar] [CrossRef]
- Ribeiro, F.R.V.; Lucena, C.A.S.; Lucinda, P.H.F. Three new Pimelodus species (Siluriformes: Pimelodidae) from the rio Tocantins drainage, Brazil. Neotrop. Ichthyol. 2008, 6, 455–464. [Google Scholar] [CrossRef]
- Fonseca, I.C.; Maciel, L.A.M.; Ribeiro, F.R.V.; Rodrigues, L.R.R. Karyotypic variation in the long-whiskered catfish Pimelodus blochii Valenciennes, 1840 (Siluriformes, Pimelodidae) from the lower Tapajós, Amazonas and Trombetas Rivers. Comp. Cytogenet. 2018, 12, 285–298. [Google Scholar] [CrossRef]
- Machado, C.B.; Ishizuka, T.K.; Freitas, P.D.D.; Valiati, V.H.; Galetti, P.M., Jr. DNA barcoding reveals taxonomic uncertainty in Salminus (Characiformes). Syst. Biodivers. 2016, 15, 372–382. [Google Scholar] [CrossRef]
- Silva-Santos, R.; Ramirez, J.L.; Galetti, P.M., Jr.; Freitas, P.D. Molecular evidences of a hidden complex scenario in Leporinus cf. friderici. Front. Genet. 2018, 9, 47. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Birindelli, J.L.; Carvalho, D.C.; Affonso, P.R.A.M.; Venere, P.C.; Ortega, H.; Carrillo-Avila, M.; Rodríguez-Pulido, J.A.; Galetti, P.M., Jr. Revealing hidden diversity of the underestimated neotropical ichthyofauna: DNA barcoding in the recently described genus Megaleporinus (Characiformes: Anostomidae). Front. Genet. 2017, 8, 149. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Santos, C.A.; Machado, C.B.; Oliveira, A.K.; Garavello, J.C.; Britski, H.A.; Galetti, P.M., Jr. Molecular phylogeny and species delimitation of the genus Schizodon (Characiformes, Anostomidae). Mol. Phylogenet. Evol. 2020, 153, 106959. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De La Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef]
- Singh, A.K. Molecular Taxonomy: Use of Modern Methods in the Identification of a Species. Minn. J. Law Sci. Technol. 2012, 2, 143–147. [Google Scholar]
- Pante, E.; Schoelinck, C.; Puillandre, N. From integrative taxonomy to species description: One step beyond. Syst. Biol. 2015, 64, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identification through DNA barcode. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Serrano, E.A.; Melo, B.F.; Freitas-Souza, D.; Oliveira, M.L.; Utsunomia, R.; Oliveira, C.; Foresti, F. Species delimitation in Neotropical fishes of the genus Characidium (Teleostei, Characiformes). Zool. Scr. 2018, 48, 69–80. [Google Scholar] [CrossRef]
- Martínez, J.G.; Rangel-Medrano, J.D.; Yepes-Acevedo, A.J.; Restrepo-Escobar, N.; Márquez, E.J. Species limits and introgression in Pimelodus from the Magdalena-Cauca River basin. Mol. Phylogenet. Evol. 2022, 173, 107517. [Google Scholar] [CrossRef] [PubMed]
- Limeira-Filho, D.; Moraes, P.S.S.; Almeida, M.S.; Silva, A.C.C.; Silva, J.L.N.; Farias-Rodrigues, L.; Teixeira, B.R.S.; Anjos, J.V.X.; Birindelli, J.L.O.; Barros, M.C.; et al. Fish diversity in the rivers that drain the Baixada Maranhense and the Mearim basin in northeastern Brazil. Braz. J. Biol. 2023, 83, e273525. [Google Scholar] [CrossRef]
- Rocha, M.S.; Zuanon, J. Pimelodidae. In Peixes do rio Madeira; Queiroz, L.J., Torrente-Vilara, G., Ohara, W.M., Pires, T.H.S., Zuanon, J., Doria, C.R.C., Eds.; Dialeto Latin American Documentary: São Paulo, Brasil, 2013; Volume 3, pp. 78–128. [Google Scholar]
- Papa, Y.; Le Bail, P.Y.; Covain, R. Genetic landscape clustering of a large DNA barcoding data set reveals shared patterns of genetic divergence among freshwater fishes of the Maroni Basin. Mol. Ecol. Resour. 2021, 21, 2109–2124. [Google Scholar] [CrossRef] [PubMed]
- Ardura, A.; Linde, A.R.; Moreira, J.C.; Garcia-Vazquez, E. DNA barcoding for conservation and management of Amazonian commercial fish. Biol. Conserv. 2010, 143, 1438–1443. [Google Scholar] [CrossRef]
- García-Dávila, C.; Castro-Ruiz, D.; Renno, J.F.; Chota-Macuyama, W.; Carvajal-Vallejos, F.M.; Sanchez, H.; Ângulo, C.; Nolorbe, C.; Alvarado, J.; Estivals, G.; et al. Using barcoding of larvae for investigating the breeding seasons of Pimelodid catfishes from the Marañon, Napo and Ucayali rivers in the Peruvian Amazon. J. Appl. Ichthyol. 2015, 31, 40–51. [Google Scholar] [CrossRef]
- Carvalho, D.C.; Oliveira, D.A.A.; Pompeu, O.S.; Leal, C.G.; Oliveira, C.; Hanner, R. Deep barcode divergence in Brazilian freshwater fishes: The case of the São Francisco River basin. Mitochondrial DNA 2011, 22, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Resende, L.C.; Carmo, A.O.D.; Núñez-Rodriguez, D.; Pimentel, J.D.S.M.; Bedore, A.G.; Leal, H.G.; Kalapothakis, E. Pimelodus maculatus (Siluriformes, Pimelodidae): Complete mtDNA sequence of an economically important fish from the São Francisco river basin. Mitochondrial DNA 2016, 1, 806–808. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.H.; Maia, G.M.; Hanner, R.; Foresti, F.; Oliveira, C. DNA barcodes discriminate freshwater fishes from the Paraíba do Sul River Basin, São Paulo, Brazil. Mitochondrial DNA 2011, 22, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Lima, T.P.C.; Egito, A.A.; Feijó, G.L.D.; Mauro, R.A.; Ferraz, A.L.J. Molecular identification and phylogenetic analysis of Siluriformes from the Paraguay River basin, Brazil. Mitochondrial DNA Part A 2016, 28, 536–543. [Google Scholar] [CrossRef]
- Pereira, L.H.G.; Hanner, R.; Foresti, F.; Oliveira, C. Can DNA barcoding accurately discriminate megadiverse Neotropical freshwater fish fauna? BMC Genet. 2013, 14, 1471–2156. [Google Scholar] [CrossRef]
- Frantine-Silva, W.; Sofia, S.H.; Orsi, M.L.; Almeida, F.S. DNA barcoding of freshwater ichthyoplankton in the Neotropics as a tool for ecological monitoring. Mol. Ecol. Resour. 2015, 15, 1226–1237. [Google Scholar] [CrossRef]
- Díaz, J.; Villanova, G.V.; Brancolini, F.; Del Pazo, F.; Posner, V.M.; Grimberg, A.; Arranz, S.E. First DNA barcode reference library for the identification of South American freshwater fish from the lower Paraná river. PLoS ONE 2016, 11, e0157419. [Google Scholar] [CrossRef]
- Calegari, B.B.; Vari, R.P.; Reis, R.E. Phylogenetic systematics of the driftwood catfishes (Siluriformes: Auchenipteridae): A combined morphological and molecular analysis. Zool. J. Linn. Soc. 2019, 187, 661–773. [Google Scholar] [CrossRef]
- Rosso, J.J.; Mabragaña, E.; Castro, G.; Díaz de Astarloa, J.M. DNA barcoding Neotropical fishes: Recent advances from the Pampa Plain, Argentina. Mol. Ecol. Resour. 2012, 12, 999–1011. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Phillos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nichlen, S.; Coulson, A.R. DNA sequencing with chaintermination inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5468. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibons, T.J. Clustal W: Improving the sensitivity of progressive multiple sequence alignment though sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kapli, P.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2017, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef]
- Pons, J.; Barraclough, T.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Valdivia, P.; Rosas-Puchuri, U.; Valdivia, N.L. SPdel: A pipeline to compare and visualize species delimitation methods for single-locus datasets. Mol. Ecol. Resour. 2023, 23, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, A.M.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree: Tree Figure Drawing Tool, Versão 1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 14 February 2023).
- Inkscape Versão 1.1. 2021. Available online: https://inkscape.org/pt-br/release/inkscape-1.1/ (accessed on 23 March 2023).
- Brown, S.D.J.; Collins, R.A.; Boyer, S.; Lefort, M.C.; Malumbres-Olarte, J.; Vink, C.J.; Cruickshank, R.H. Spider: An R package for the analysis of species identity and evolution, with particular reference to DNA barcoding. Mol. Ecol. Resour. 2012, 12, 562–565. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing, Versão 4.3.1. R Foundation for Statistical Computing: Vienna, Austria, 2023. Available online: https://www.r-project.org/ (accessed on 10 May 2023).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of Birds through DNA Barcodes. PLoS Biol. 2004, 10, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: Na approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Garavello, J.C.; Ramirez, J.L.; Oliveira, A.K.D.; Britski, H.A.; Birindelli, J.L.; Galetti, P.M., Jr. Integrative taxonomy reveals a new species of Neotropical headstanding fish in genus Schizodon (Characiformes: Anostomidae). Neotrop. Ichthyol. 2021, 19, e210016. [Google Scholar] [CrossRef]
- Machado, C.B.; Galetti, P.M., Jr.; Carnaval, A.C. Bayesian analyses detect a history of both vicariance and geodispersal in Neotropical freshwater fishes. J. Biogeogr. 2018, 45, 1313–1325. [Google Scholar] [CrossRef]
- Lima, F.C. Revision of the smaller-sized dorados (Salminus), with comments on the monophyly of the genus and its biogeography (Characiformes: Bryconidae). Zootaxa 2022, 5226, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, G.J.; Rodriguez, M.S.; Roxo, F.F.; Foresti, F.; Oliveira, C. Using different methods to access the difficult task of delimiting species in a complex Neotropical hyperdiverse group. PLoS ONE 2015, 10, e0135075. [Google Scholar] [CrossRef]
- Rossini, B.C.; Oliveira, C.A.M.; Melo, F.A.G.D.; Bertaco, V.D.A.; Díaz de Astarloa, J.M.; Rosso, J.J.; Foresti, F.; Oliveira, C. Highlighting Astyanax species diversity through DNA barcoding. PLoS ONE 2016, 11, e0167203. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Galetti, P.M., Jr. DNA barcode and evolutionary relationship within Laemolyta Cope 1872 (Characiformes: Anostomidae) through molecular analyses. Mol. Phylogenet. Evol. 2015, 93, 77–82. [Google Scholar] [CrossRef]
- Nascimento, M.H.S.; Aragão, D.G.; Silva, J.L.N.; Lima, R.C.; Birindelli, J.L.O.; Fraga, E.C.; Barros, M.C. The DNA barcode reveals cryptic diversity and a new record for the genus Leporinus (Characiformes, Anostomidae) in the hydrographic basins of central northern Brazil. PeerJ 2023, 11, e15184. [Google Scholar] [CrossRef]
- Melo, B.F.; Dorini, B.F.; Foresti, F.; Oliveira, C. Little divergence among mitochondrial lineages of Prochilodus (Teleostei, Characiformes). Front. Genet. 2018, 9, 107. [Google Scholar] [CrossRef]
- Arruda, P.S.; Ferreira, D.C.; Oliveira, C.; Venere, P.C. DNA Barcoding reveals high levels of divergence among mitochondrial lineages of Brycon (Characiformes, Bryconidae). Genes 2019, 10, 639. [Google Scholar] [CrossRef]
- Rocha, M.S.; Pavanelli, C.S. Taxonomy of Pimelodus brevis Marini, Nichols & La Monte, 1933 (Siluriformes: Pimelodidae), an uncertain species from the rio Paraná basin. Neotrop. Ichthyol. 2014, 12, 755–760. [Google Scholar] [CrossRef]
- Britski, H.A.; Silimon, K.Z.S.; Lopes, B.S. Peixes do Pantanal: Manual de Identificação, 2nd ed.; EMPRAPA: Brasília, Brazil, 2007. [Google Scholar]
- Almeida, F.S.D.; Sodré, L.M.K.; Contel, E.P.B. Population structure analysis of Pimelodus maculatus (Pisces, Siluriformes) from the Tietê and Paranapanema rivers (Brazil). Genet. Mol. Biol. 2003, 26, 301–305. [Google Scholar] [CrossRef]
- Garcia, C.; Moreira Filho, O. Cytogenetical analyses in three fish species of the genus Pimelodus (Siluriformes: Pimelodidae) from rio São Francisco: Considerations about the karyotypical evolution in the genus. Neotrop. Ichthyol. 2005, 3, 285–290. [Google Scholar] [CrossRef]
- Britski, H.A.; Silimon, K.Z.S.; Lopes, B.S. Peixes do Pantanal: Manual de Identificação, 1st ed.; Embrapa: Corumbá, Brazil, 1999. [Google Scholar]
- Eigenmann, C.H. The freshwater fishes of British Guiana, including a study of the ecological grouping of species and the relation of the fauna of the plateau to that of the lowlands. Mem. Carnegie Museum. Carnegie Inst. 1912, 5, 171–177. [Google Scholar] [CrossRef]
- Abreu, J.M.S.; Craig, J.M.; Albert, J.S.; Piorski, M.N. Historical biogeography of fishes from coastal basins of Maranhão State, northeastern Brazil. Neotrop. Ichthyol. 2019, 17, e180156. [Google Scholar] [CrossRef]
- Abreu, J.M.S.; Saraiva, A.C.S.; Albert, J.S.; Piorski, N.M. Paleogeographic influences on freshwater fish distributions in northeastern Brazil. J. South Am. Earth Sci. 2020, 102, 102692. [Google Scholar] [CrossRef]
- Ramos, T.P.A.; Ramos, R.T.C.; Ramos, S.A.Q.A. Ichthyofauna of the Parnaíba river Basin, Northeastern Brazil. Biota Neotrop. 2014, 14, e20130039. [Google Scholar] [CrossRef]
- Silva, M.J.; Costa, B.G.; Ramos, T.P.A.; Auricchio, P.; Lima, S.M.Q. Ichthyofauna of the Gurgueia river, Parnaíba river basin, northeastern Brazil. Check List 2015, 11, 1765. [Google Scholar] [CrossRef]
- Silva, S.C.D.; Medeiros, L.S.D.; Bezerra Neto, M.F.; Silva, M.J.D.; Ramos, T.P.A.; Lima, F.C.T.D.; Lima, S.M.Q. Ichthyofauna of the Nascentes do Rio Parnaíba National Park: Watershed divide in the main agricultural encroachment area in the Brazilian Cerrado. Biota Neotrop. 2023, 23, e20221414. [Google Scholar] [CrossRef]
- Rosa, R.S.; Menezes, N.A.; Britski, H.A.; Costa, W.J.E.; Groth, F. Diversidade, padrões de distribuição e conservação dos peixes da caatinga. In Ecologia e Conservação da Caatinga; Leal, I.R., Tabarelli, M., Silva, J.M.C., Eds.; Universitária da Universidade Federal de Pernambuco: Recife, Brasil, 2003; pp. 135–162. [Google Scholar]
- Hubert, N.; Renno, J.F. Historical biogeography of South American freshwater fishes. J. Biogeogr. 2006, 33, 1414–1436. [Google Scholar] [CrossRef]
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. Bioscience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Ribeiro, M.F.R.; Piorski, N.M.; Almeida, Z.D.S.D.; Nunes, J.L.S. Fish aggregating known as moita, an artisanal fishing technique performed in the Munim River, State of Maranhão, Brazil. Bol. Inst. Pesca 2014, 40, 677–682. [Google Scholar]
- Matavelli, R.; Campos, A.M.; Vale, J.; Piorski, N.M.; Santos Pompeu, P. Ichthyofauna sampled with tadpoles in northeastern Maranhão state, Brazil. Check List 2015, 11, 1550. [Google Scholar] [CrossRef]
- Nunes, J.L.S.; Sousa Costa, M.; Rêgo Ribeiro, M.F.; Costa, M.S.; Bonfim, M. Length–weight relationship for freshwater fish species from Brazilian Cerrado. J. Appl. Ichthyol. 2019, 35, 1047–1049. [Google Scholar] [CrossRef]
- Brito, P.S.D.; Guimarães, E.C.; Ferreira, B.R.A.; Ottoni, F.P.; Piorski, N.M. Freshwater fishes of the Parque Nacional dos Lençóis Maranhenses and adjacent areas. Biota Neotrop. 2019, 19, e20180660. [Google Scholar] [CrossRef]
- Oliveira, E.D.S.; Guimarães, E.C.; Brito, P.S.D.; Vieira, L.D.O.; Oliveira, R.F.D.; Campos, D.S.; Katz, A.M.; South, J.; Nunes, J.L.S.; Ottoni, F.P. Ichthyofauna of the Mata de Itamacaoca, an urban protected area from the upper Munim River basin, Northern Brazilian Cerrado. Biota Neotrop. 2020, 20, e20201116. [Google Scholar] [CrossRef]
- Vieira, O.L.; Campos, D.S.; Oliveira, R.F.; South, J.; Coelho, M.S.P.; Paiva, M.J.S.; Bragança, P.H.N.; Guimarães, E.C.; Katz, A.M.; Brito, O.S.; et al. Checklist of the fish fauna of the Munim River Basin, Maranhão, north-eastern Brazil. Biodivers. Data J. 2023, 11, e98632. [Google Scholar] [CrossRef]
- Mees, G.F. The Auchenipteridae and Pimelodidae of Suriname (Pisces, Nematognathi). Zool. Verh. 1974, 132, 1–246. [Google Scholar]
- Rocha, M.S.; Ribeiro, F.R.V. A new species of Pimelodus LaCépède, 1803 (Siluriformes: Pimelodidae) from rio Itacaiunas, rio Tocantins basin, Brazil. Zootaxa 2010, 2343, 57–65. [Google Scholar] [CrossRef]
- Lucinda, P.H.F.; Ribeiro, F.R.V.; Lucena, C.A.S. Pimelodus quadratus, a new long-whiskered catfish from the rio Tocantins drainage, Brazil (Siluriformes: Pimelodidae). Ichthyol. Explor. Freshw. 2016, 27, 337–345. [Google Scholar]
- Ribeiro, F.R.V.; Lucena, C.A.S. Nova espécie de Pimelodus (Siluriformes, Pimelodidae) dos rios Tapajós e Tocantins, Brasil. Iheringia. Porto Alegre. Iheringia Ser. Zool. 2006, 96, 321–327. [Google Scholar] [CrossRef]






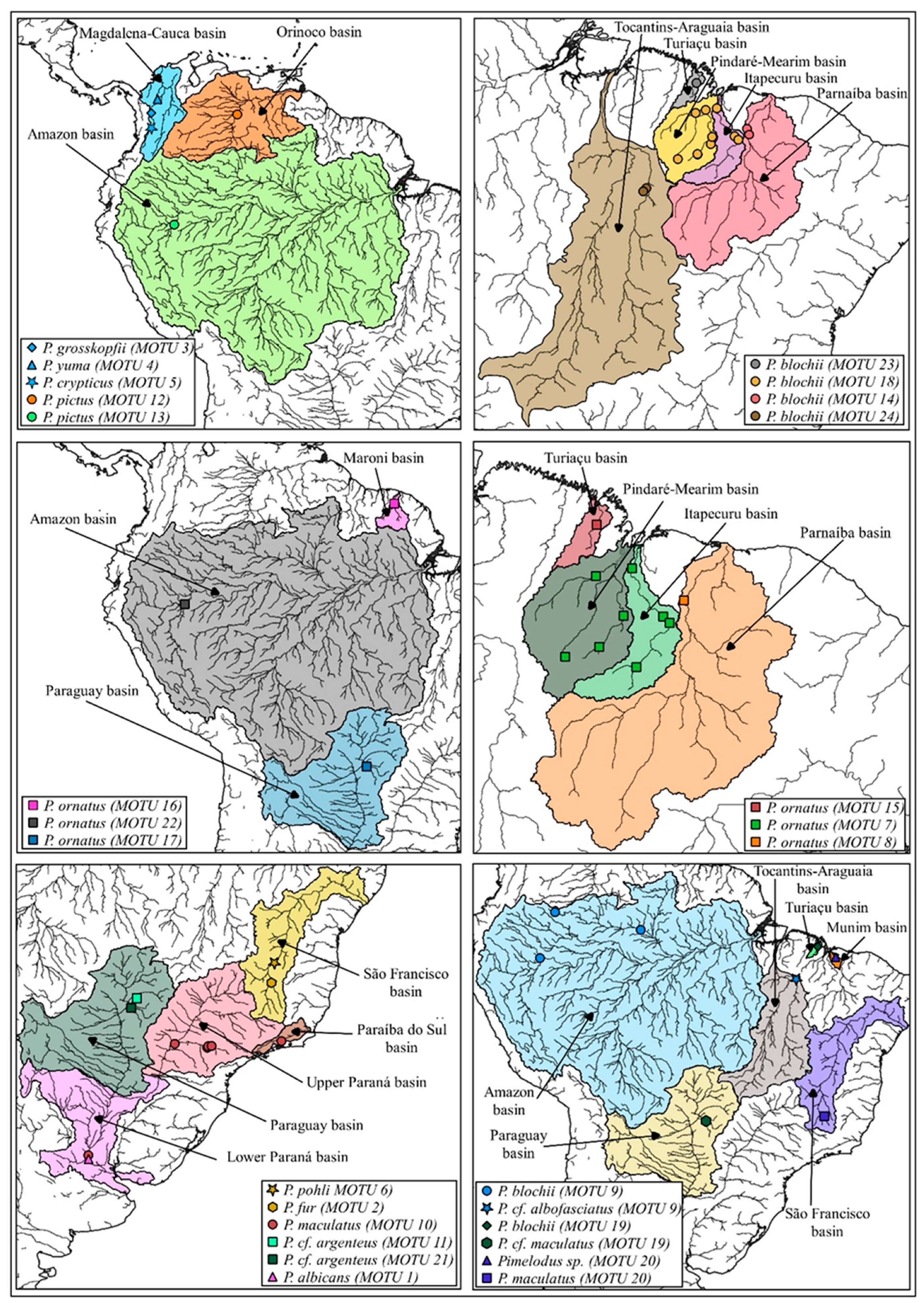{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Intra- | Maximum Intra- | NN | Distance to NN | |
|---|---|---|---|---|
| Nominal Species | ||||
| P. albofasciatus | 0.00 | 0.00 | P. blochii | 0.00 |
| P. albicans | 0.00 | 0.00 | Pimelodus sp. | 2.84 |
| P. argenteus | 2.11 | 3.01 | P. blochii | 0.81 |
| P. blochii | 1.19 | 3.53 | P. maculatus/P. albofasciatus | 0.00 |
| P. crypticus | 0.07 | 0.22 | P. albicans | 6.19 |
| P. fur | 0.00 | 0.00 | P. albofasciatus | 3.51 |
| P. grosskopfii | 0.00 | 0.00 | P. crypticus | 6.95 |
| P. maculatus | 2.17 | 7.15 | P. blochii/Pimelodus sp. | 0.00 |
| P. ornatus | 2.89 | 6.17 | Pimelodus sp. | 13.1 |
| P. pictus | 2.47 | 4.97 | P. pohli | 9.66 |
| P. pohli | 0.49 | 0.98 | P. albofasciatus | 2.83 |
| Pimelodus sp. | 0.28 | 0.46 | P. maculatus | 0.00 |
| P. yuma | 0.00 | 0.00 | P. maculatus/P. blochii | 4.48 |
| Consensus MOTUs | ||||
| P. albicans (MOTU 1) | 0.00 | 0.00 | Pimelodus sp./P. maculatus MOTU 20 | 2.84 |
| P. fur (MOTU 2) | 0.00 | 0.00 | P. blochii/P. cf. albofasciatus MOTU 09 | 3.51 |
| P. grosskopfii (MOTU 3) | 0.00 | 0.00 | P. crypticus MOTU 05 | 6.95 |
| P. yuma (MOTU 4) | 0.00 | 0.00 | P. blochii/P. cf. maculatus MOTU 19 | 4.48 |
| P. crypticus (MOTU 5) | 0.07 | 0.22 | P. albicans MOTU 01 | 6.19 |
| P. pohli (MOTU 6) | 0.49 | 0.98 | P. blochii/P. cf. albofasciatus MOTU 09 | 2.83 |
| P. ornatus (MOTU 7) | 0.07 | 0.15 | P. ornatus MOTU 08 | 2.91 |
| P. ornatus (MOTU 8) | - | - | P. ornatus MOTU 07 | 2.91 |
| P. blochii/P. cf. albofasciatus (MOTU 9) | 0.07 | 0.33 | P. blochii/P. cf. maculatus MOTU 19 | 1.18 |
| P. maculatus (MOTU 10) | 0.20 | 0.98 | Pimelodus sp./P. maculatus MOTU 20 | 1.14 |
| P. cf. argenteus (MOTU 11) | - | - | P. blochii MOTU 24 | 0.81 |
| P. pictus (MOTU 12) | 0.30 | 0.45 | P. pictus MOTU 13 | 4.47 |
| P. pictus (MOTU 13) | - | - | P. pictus MOTU 12 | 4.47 |
| P. blochii (MOTU 14) | 0.10 | 0.31 | P. blochii MOTU 18 | 0.78 |
| P. ornatus (MOTU 15) | 0.02 | 0.15 | P. ornatus MOTU 22 | 0.79 |
| P. ornatus (MOTU 16) | 0.09 | 0.31 | P. ornatus MOTU 17 | 2.83 |
| P. ornatus (MOTU 17) | 0.32 | 0.65 | P. ornatus MOTU 15 | 1.48 |
| P. blochii (MOTU 18) | 0.05 | 0.31 | P. blochii MOTU 14 | 0.78 |
| P. blochii/P. cf. maculatus (MOTU 19) | 0.10 | 0.49 | P. blochii/P. cf. albofasciatus MOTU 09 | 1.18 |
| Pimelodus sp./P. maculatus (MOTU 20) | 0.27 | 0.93 | P. maculatus MOTU 10 | 1.14 |
| P. cf. argenteus (MOTU 21) | 0.49 | 0.49 | P. blochii/P. cf. albofasciatus MOTU 09 | 1.65 |
| P. ornatus (MOTU 22) | - | - | P. ornatus MOTU 15 | 0.79 |
| P. blochii (MOTU 23) | 0.42 | 0.95 | P. blochii MOTU 24 | 0.78 |
| P. blochii (MOTU 24) | 0.00 | 0.00 | P. blochii MOTU 23 | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limeira Filho, D.; França, E.R.d.R.; Costa, D.K.d.P.; Lima, R.C.; Nascimento, M.H.S.d.; Batista, J.d.S.; Barros, M.C.; Fraga, E.d.C. Molecular Evidence Reveals Taxonomic Uncertainties and Cryptic Diversity in the Neotropical Catfish of the Genus Pimelodus (Siluriformes: Pimelodidae). Biology 2024, 13, 162. https://doi.org/10.3390/biology13030162
Limeira Filho D, França ERdR, Costa DKdP, Lima RC, Nascimento MHSd, Batista JdS, Barros MC, Fraga EdC. Molecular Evidence Reveals Taxonomic Uncertainties and Cryptic Diversity in the Neotropical Catfish of the Genus Pimelodus (Siluriformes: Pimelodidae). Biology. 2024; 13(3):162. https://doi.org/10.3390/biology13030162
Chicago/Turabian StyleLimeira Filho, Daniel, Elidy Rayane de Rezende França, Dalton Kaynnan de Prado Costa, Renato Correia Lima, Maria Histelle Sousa do Nascimento, Jacqueline da Silva Batista, Maria Claudene Barros, and Elmary da Costa Fraga. 2024. "Molecular Evidence Reveals Taxonomic Uncertainties and Cryptic Diversity in the Neotropical Catfish of the Genus Pimelodus (Siluriformes: Pimelodidae)" Biology 13, no. 3: 162. https://doi.org/10.3390/biology13030162